Over de Zaadvorming bij den Dar.
Inleiding.
Dat het normale verloop van de sperma (zaad) vorming bij de honingbij-dar eigenlijk een afwijkende vorm is van het algemeene grondplan der Spermatogenese (zaad- of spermavorming) bij de insekten, maakt dit onderwerp tot één der merkwaardigste hoofdstukken uit de biologie (levensleer), speciaal der voortplanting, van onze honingbij.
Kennis is macht.
Voor imker-koninginnetelers zijn de feiten niet slechts interessant, maar behoort de nauwkeurige kennis de grondslag te zijn om de belangrijkste factoren uit de erfelijkheidsleer (Genetica) te begrijpen, welke leer hij tevens tot in kleinste kleinigheden moet kennen. Nu er ook in Nederland in breeder lagen wat belangstelling komt voor koninginne(n)teelt, wensch ik er van den beginne af duidelijk op te wijzen, dat, zooals er tusschen bijenhouderij en bijenteelt een enorm verschil bestaat, een zelfde groot verschil eveneens bestaat tusschen koninginnenteelt als produceeren van veel moeren en koninginneteelt als fokken van edele-rasmoeren. Voor de eerste is slechts wat praktische kennis noodig van de materialen en van het hoe te doen; de tweede stelt behalve aan de technische vaardigheid hooge eischen aan de kennis van morphologie (uitwendige vormkennis) en van de anatomie (inwendige vormkennis), en zooals gezegd van de erfelijkheidsleer, dat hij de biologie van de bij moet kennen en een prima imker moet zijn is wel vanzelfsprekend, terwijl een aangeboren, fijne intuïtie noodzakelijk is om tot een werkelijke fokker of teler te zijn geroepen, met alle moreele verplichtingen van dien.
Belang aangetoond.
Na aldus de nood. zakelijkheid der preciese kennis der onderdeelen van de bijenteelt te hebben aangetoond en daar mede het opnemen van deze artikelen in on Maandschrift te hebben gerechtvaardigd, immers er zal langzamerhand ook wat in opgenomen wor den voor meer gevorderde en ontwikkelder imkers zoo zal dan dit opstel het eerste zijn als inleiding tot de reeks over de erfelijkheidsleer in de bijenteelt.
Orgaan en tijd.
Zooals bij de ontwikkeling van het ei nader zal worden aangetoond, is het geslacht van het Imago (volwassen insekt), dus of het een dar of een koningin, ev. werkster is, bepaald op het oogenblik dat het ei gelegd wordt. Wanneer aan het einde van de derde eidag het larfje, in dit opstel dus het darrenlarfje, de eischaal doorbreekt om eerste dags larf te worden van 0.35 milligram en 1.2 m.m. grootte dan is dit larfje reeds in het bezit van een paar duidelijke zaad- of teeltballen (Testes) waarin de aanleg van de zaadbuisjes (Testiolen of Follikels) al te onderscheiden zijn.
Het is in deze Follikels dat de spermiogenese (zaadvorming) zich voltrekt.
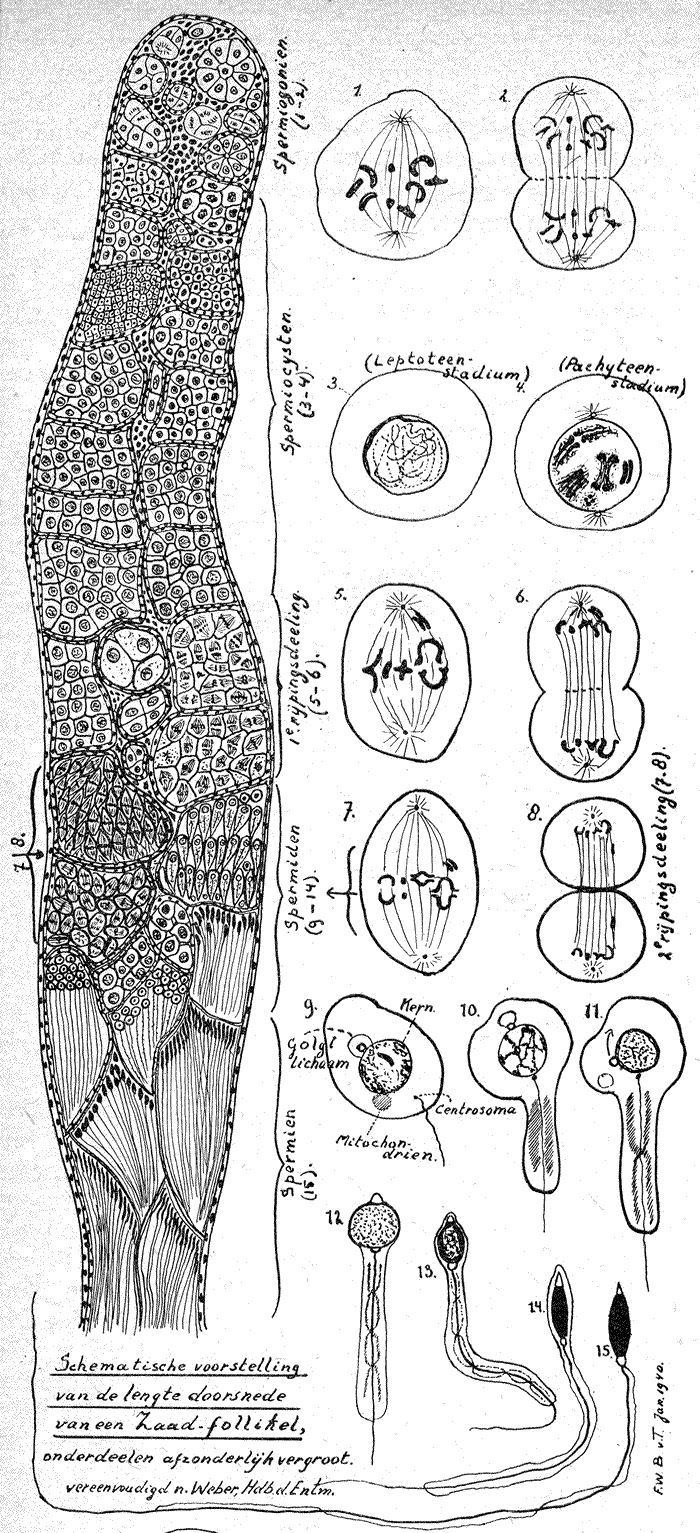
De Oergeslachtscellen (Oerspermiogonien) worden bij de honingbij namelijk reeds eenige uren na het leggen van het ei gevormd door afzondering van enkele kernen der eerste kerndeelingen, wat bij het mannelijke- of darren-ei (al klinkt het wat eigenaardig), waarbij geen bevruchting plaats heeft, dus ook niet behoeft te wachten tot de versmelting van zaadkern met eikern heeft plaats gehad. Normaal heeft deze Blastos (afzondering der oergeslachtscellen, die dan door een geheimzinnige kracht door de eidooier heen trekken naar de plek waar later de testes liggen) dus zelfs reeds plaats voor de aanleg van een Blastoderm (eerste cellaag aan de oppervlakte der eidooier). De teeltballen worden gevormd of zijn opgebouwd uit een groot aantal gekronkelde zaadbuisjes, ongeveer 200 per bal bij de honingbij-dar, welke door een omhulsel samen gehouden worden, waarover t.z.t. uitvoeriger.
Zooals reeds opgemerkt is heeft in deze zaadbuisjes de vorming plaats van het mannelijk geslachtsprodukt de Spermien of Spermatozoën of Spermatozoïden, als geheel ook genoemd het Sperma. Dit proces der zaadvorming (spermiogenese) is bij de dar aan het einde van de vijfde poptoestand-dag (dus ± 22 dagen na het leggen van het ei of ± 2 dagen voordat de dar uitkomt) geheel voltooid en afgesloten.
Normale verloop bij de insekten.²)
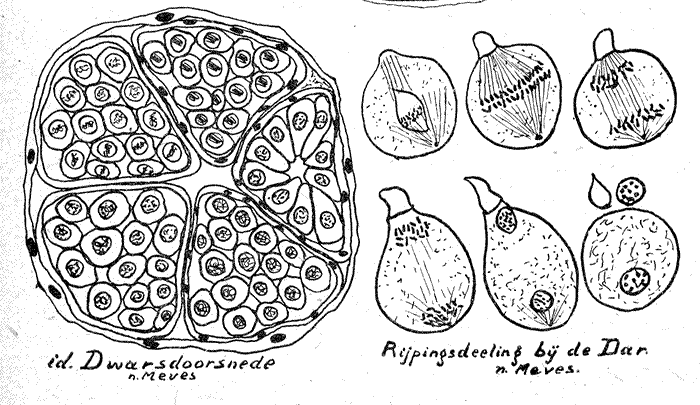
Aan de hand van bijgaande schetsteekening zal het nu niet moeilijk meer zijn het ontstaan der Spermien zelve te begrijpen. De geheele Follikel bestaat uit een vliezig eencellaag dik omhulsel en de inhoud welke door epitheliumachtig gelagerde cellen (Cystencellen) in afzonderlijke vakjes (Spermiocysten = Spermatocysten) wordt verdeeld, welke dicht opeengedrongen tot aan de opening van de Follikels liggen.
Het gesloten einde van de Follikels heet Apikale einde. In dit apikale einde liggen de mannelijke oergeslachtscellen (oerspermiogonien), omgeven door een groot aantal zgn. somatische cellen, hier genoemd Follikelcellen. Bij sommige insekten treft men één buitengewoon groote Oergeslachtscel (de Apikaalcel) aan, die echter niet aan de zaadvorming deelneemt, maar vermoedelijk als voedingscel fungeert. Terwijl nu in dit apikale einde ook wel genoemd Germarium, de spermiogonien of permatogonien, die zich door deeling vermenigvuldigd hebben, steeds verder naar de opening toe zakken, omgeven de somatische cellen hen, als het ware met een celvliesomhulsel, dat dan later de wand vormt om de zich vormende Cyste, in dit geval de Spermiocyste. Sommige dezer cysten kunnen beduidend vergroot zijn, maar zij geven geen spermien, schijnen evenals de apikaalcel voor de voeding te dienen.
Iedere Cyste bevat dus de door de gewone mitotische (indirekte) deeling ontstane afstammelingen van één Oergeslachtscel.
In het stadium der eerste deelingen is het mogelijk de voor de erfelijkheid zoo uiterst gewichtige Chromosomen en hun aantal (het Genom) vast te stellen.
De vorming.
De Spermiogenese wordt in twee phasen onderverdeeld, namelijk die der zaadcelvorming of Spermiocytogenese,A.; en die der zaadvormgeving
of Spermiohistogenese,B.
(men vergelijke bijgaande schematische schetsen).
A. De Spermiocytogenese.
Dit is de periode vanaf de vorming der Oergeslachtscellen tot en met de rijpingsdeelingen. Hierin onderscheiden wij twee afzonderlijke phasen:
1. die der vermenigvuldiging en
2. die der rijpingsdeelingen.
1. De phase der vermenigvuldiging.
De primaire, nog niet als Cysten ingesloten, spermiogonien, deelen zich mitotisch (ook wel genoemd Karyokinetisch) zes à acht keer, dan ontstaan de volgende eveneens mitotische aekuatische (gelijkgedeelde) deelingen, welke zich op de voren beschreven wijze met een huidje van somatische cellen omgeven en zoo tot eigenlijke Cysten worden. Het typische diploide (man- plus vrouwlijk) chromosomen getal blijft in het normale insektengeval dus nog steeds behouden, bij de dar is dit zooals in de inleiding reeds werd opgemerkt iets anders, wat nog afzonderlijk beschreven zal worden. De vermenigvuldigingsdeelingen verschillen van de gewone slechts door een sneller verloop, wat hierdoor tot stand komt, dat de celkernen tusschen de deelingen in niet geheel tot het ruststadium terugkeeren en de dochtercellen vóór nieuwe deelingen, niet in alle gevallen, eerst tot de grootte van de moedercel uitgroeien.
Bij de spermiocysten, die evenwel wèl uitgroeien, keert de kern ook niet tot een ruststadium terug, de chromosomen lossen zich tot chromatine korreltjes op, maar deze ordenen zich tot een kluwen dunne draadjes van het diploide chromosomen getal (Leptoteenstadium). Deze draadjes vertoonen kort daarop een lengtekloof (Diploteenstadium), daarna worden zij dikker en korter (Pachyteenstadium).
Daar waar het tellen van diploteen- en pachyteendraadjes mogelijk is, vertoont zich merkwaardigerwijze slechts het halve aantal van de oorspronkelijke chromosomen. Er heeft een pseudoreductie plaats gehad, doordat de chromatinedraadjes zich soortgelijkelijk paarsgewijze samen gevoegd hebben (Konjugatie of Syndese) en zoo tot dubbelchromosomen zijn geworden en Dyaden genoemd worden. Daar evenwel ieder draadje in het diploteenstadium een lengtespleet vertoonde, zijn eigenlijk vierlingen of Tetraden ontstaan.
2. De phase der rijpingsdeelingen. (Meiosis).
Na deze vermenigvuldigingsdeelingen volgen nu nog twee deelingen, de rijpingsdeelingen, welke zich daardoor kenmerken, dat het chromosomengetal (gewoonlijk bij de eerste deeling) tot de helft wordt teruggebracht. Hierbij ontstaan dan uit één diploide spermiocyste eerst twee haploide praespermiden en dan vier haploide spermiden.
In normale gevallen (het erochromomosie komt misschien bij de parthenogenes nog eens ter sprake) verloopt de reductiedeeling als volgt zoo, dat bij de eerste deeling iedere Tetrade in de twee deelen uiteenvalt die zich eerst door de Syndese hadden verbonden. Ieder dezer helften, dus ieder een oorspronkelijk chromosoom, treedt nu in één der beide dochtercellen, in dit geval de prae-spermiden.
Bij de tweede reductiedeeling splijt iedere tetradehelft dan volgens de lengtespleet in tweeën volgens de gewone aequatische karyokinese. Hoe het verder met deze chromosomen gesteld is, zal bij de erfelijkheidsleer uitvoerig behandeld worden.
B. De Spermiohistogenese.
Deze omvat de periode waarin de ronde spermiden uitgroeien en tot gestaarte spermiden, de voltooide zaaddiertjes. Bijgaande sterk schematische en vereenvoudigde teekeningen toonen duidelijk wat hier beschreven is. Het verloop betreft wel voornamelijk de kern, waarin de oplossing van het chromatine, dat bijna onkleurbaar wordt, opvalt. Dan strekt zich de kern, wordt weer extra kleurbaar en vormt de hoofdmassa van de spermienkop. Het heldere spitsje ontstaat waarschijnlijk uit het zgn. Golgiapparaat. Terwijl vermoedelijk het middenstuk door het Centrosoma (Centriool) wordt gevormd, waaruit het staartje groeit, oorspronkelijk nog door een trilzoom omgeven, welke verdwijnt.
Hoe het bij de Dar anders is.
De van het normale verloop der Spermiogenese afwijkende vorm bij de honingbij-dar ontstaat daar- door, dat de dar die zelf ontstaan is uit een onbevrucht ei (parthenogenetisch) slechts het halve aantal chrosomen in de cellen bezit. Zou hier dus ook de reductiedeeling plaats hebben, dan zou het constante chromosomen getal (Het Genom) verloren gaan. Daarom heeft bij de dar een schijnreductie plaats, die wel de massa, maar niet het aantal der chromosomen vermindert en welke zich als volgt voltrekt. In het pachyteenstadium snoert eerst een stukje van de cel zich zonder kern af, daarna ordenen zich de 16 chromosomen (er komen bij de dar ook wel 8 dubbelchromosomen voor) in het midden van de cel. Deze worden allen langs gespleten, de eene helft blijft als nieuwe kern in de oude cel welke tot spermatozoide uitgroeit en de andere helft vormt een zich vergrootende kern zonder cel, welke uitgestooten wordt. Wat met de uitgestooten stukken gebeurt is niet bekend.
Het aantal.
Alle spermien liggen bij de dar gelijkgericht (unipolair), de koppen allen één richting uit. Als alle spermatozoën of spermatozoiden voltooid zijn heeft de dar er ongeveer 200.000.000
geproduceerd, en hoewel ieder spermatozoide slechts 275 mikra (1 mikron is 0.001 m.m.) groot is, zouden de zaaddiertjes van één dar aan elkaar gelegd toch een lengte van 55 k.m. halen.
Hoe het verder gaat.
Hoe het verder gaat als deze spermatozoën tot een zaadpatroon vereenigd worden, is een volgend hoofdstuk. Dat zij soms tot bundels vereenigd zijn is beschreven in een opstel: Spermiozeugmen en Darrenbroedigheid.
LAREN N.H.,
F.W. BEEKHUIS VAN TIL.
1) Opzettelijk zijn in dit opstel de termini technisi, d.i. de technische- of vaktermen en hun verklaringen opgenomen en om er mede vertrouwd te raken zijn zij vaak herhaald; voor het goed kunnen lezen van vakliteratuur is dit onvermijdelijk.
2) Uitvoerig beschreven door Depdolla in Schröders Hndb. d. Entm. De in dit opstel vereenvoudigde en beknopte beschrijving volgt in hoofdlijnen die uit Weber, H. Lhrb. d. Entm. 1933, gecomb. met Zander en Meves.